β-内酰胺酶的主要功能
发布日期:2020/10/18 17:56:53
背景及概述[1]
能裂解青毒素和头孢菌素类抗生素的β-内酰胺环,使它们灭活的水解酶,称为β-内酰胺酶。以青霉素为底物的称为青霉素酶,以头孢菌素作为底物的称头孢菌素酶。根据β-内酰胺酶的基质特异性,可大致分为青霉素酶、头孢菌毒酶、肟型头孢菌素酶三类。青霉素酶易于分解青霉素类抗生素,头孢菌素酶对分解头孢菌素类抗生素的活性较高,肟型头胞菌素酶则对青霉素和头孢菌素类都有分解作用,特别更易予分解肟型头孢菌素。产生青霉素酶的遗传因子,主要基点在胞质体DNA上,而产生头孢菌素酶和肟型头孢菌素的遗传因子,则大都在宿主染色体上。青霉素酶从其基特异性或酶的生化性状上的特点,又可分为5种类型(即青霉素酶Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ型)。头孢菌素酶和肟型头孢菌素酶属于菌株特有的酶,上述这些酶的精制品、用兔做免疫试验均可得到抗体。然而,各种类型的β-内酰胺酶,在各自特有的免疫学方面,却无相互交叉的现象。
分子结构[2]
目前,β-内酰胺酶的氨基酸序列、立体结构、功能及三者之间的相互关系已基本明确。β-内酰胺酶与D-丙氨酰-D-丙氨酸转肽酶的三维结构相似,二者来源于同一酶家族。不同的β-内酰胺酶,其分子一级结构迥异,但三级结构相似。依据Ambler分类法,B类酶即金属β-内酰胺酶的活性位点几乎都有金属离子Zn2+;B类酶之外的其他类酶,统称丝氨酸β-内酰胺酶,其活性位点都有丝氨酸(Ser70)。
丝氨酸β-内酰胺酶在参与底物识别和催化的过程中,均依赖若干高度保守的序列,以一定空间结构协同发挥作用。Ω环作为特别的联体,可把亲核水解反应所需的水分子准确定位。A类酶中第224位点的精氨酸则可同时定位另一个水分子,此双水分子同时定位的结构可水解克拉维酸和舒巴坦等;而C类酶无此结构,故不能水解克拉维酸。Ω环高度保守的第166位点的谷氨酸可促进β-内酰胺酶脱酰基。Ω环与其他联体各自作为一个结构域单元,它们之间的相对位置所形成的空腔,即为β-内酰胺酶与底物作用的结合腔。结合腔中央是酶的活性位点Ser70。Ser70的-OH、-NH-与237位点的丙氨酸残基的-NH-共同构成“氧离子口袋”,使抗生素β-内酰胺环上的羰基极化,形成酰化酶复合物。β-内酰胺酶的活性部位结构,不论是源于染色体还是质粒,都有一个α螺旋结构域与5条反向平行的β片层结构,因此,结合腔与“氧离子口袋”的位置关系恰是α螺旋结构域的外层与内层的关系,该α螺旋结构域其实更像是“@”形的结构。通过氨基酸的替换使结合腔的结构改变,从而具有较大空间位阻的底物可到达活性位点与Ser70结合,这是细菌对抗生素耐药及耐药谱扩大的重要机制。金属β-内酰胺酶已被证实的部分酶为晶体结构,其结构主要相似之处为:都有与Zn+结合的保守系列HXHXD区,即116~120位点氨基酸;均有一个同样的αββα的夹心结构,其外侧为两个α螺旋,中心为β片层,β片层一端含有Zn+结合位点。因所含金属离子数目各不相等,形成了酶与底物结合方式的多样化。该类酶主要是通过金属离子与氨基酸形成配位化合物以干预活性位点的反应。而活性位点与Zn+亲和力的强弱不一,这是各金属酶水解能力互异的重要原因。
分类[2]
总体上分为活性位点含丝氨酸残基的丝氨酸β-内酰胺酶与活性依赖金属离子(多为Zn2+)的金属β-内酰胺酶。该分类方法有助于临床对酶抑制剂的选择。1995年Bush-Jacoby-Medeiros(B-J-M)分类法形成并被普遍接受。该法根据抑制剂、物及结构的不同,把β-内酰胺酶划分为4群。与此同时,根据氨基酸与核苷酸序列的不同,各群又分属于相应的分子类型。
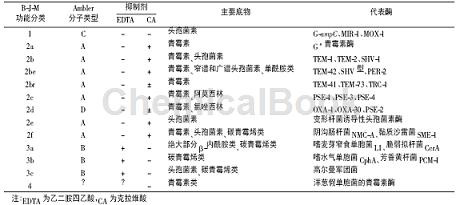
第1群大多为染色体介导,少数由质粒介导,均属于不被克拉维酸抑制的头孢菌素酶。第2群由8个亚群组成,可被β-内酰胺酶抑制剂抑制,其中,少数不被克拉维酸抑制。该群酶的种类和数量较多,多为质粒介导。第3群由3个亚群组成,其活性依赖金属离子(多为Zn2+)的参与,因此又称金属β-内酰胺酶。该群酶可使几乎所有的β-内酰胺类抗生素失活,如2010年引起全球广泛关注的NewDelhimetallo-β-lactamases-1(NDM-1)就是一个耐药性超强的金属β-内酰胺酶,故该群酶临床上需要特别注意鉴别。随着NDM-1超级细菌的发现,研究者逐渐将目光更多地投向金属β-内酰胺酶的研究,该群酶对氨曲南敏感,可被乙二胺四乙酸(EDTA)、菲咯啉及巯基化合物抑制。第4群为青霉素酶,不为克拉维酸所抑制,分子类别待定,并尚不清楚是否被EDTA抑制。
功能[3]
β-内酰胺酶对抗生素的作用主要有水解和非水解两种方式[4]。大多数β-内酰胺酶的活性位点具有一个纵行沟状结构,该结构疏松易弯曲,利于底物的结合。抗生素β-内酰胺环上的羰基碳可以不可逆的结合在该活性位点处的丝氨酸上,使抗生素β-内酰胺环解开,造成抗生素降解,避免抗生素的检出。另外一些抗生素如金属酶,它们是利用二价金属离子与组氨酸或半胱氨酸结合,并与抗生素羰基中的酰胺键相互作用,抑制抗生素发挥作用,避免抗生素的检出。目前国际上研究比较活跃的是超广谱β-内酰胺酶和头孢菌素酶。
耐药机制[2]
抗生素是对细菌的生存构成的选择性压力之一,而细菌耐药则是对该压力做出的自然选择。在该压力作用下,少数细菌成为“持留菌”并继续生长繁殖,“持留菌”被认为是当前细菌感染治疗无效的主要因素。新近研究逐步揭示了“持留菌”耐药机制,即“蛋白网络”理论的提出,有望使临床耐药菌感染的治疗获得突破性进展。β-内酰胺酶的产生与自然界微生物生成的β-内酰胺类抗生素的刺激有关,与之相关的抗生素耐药性继而产生,因此在发现和使用β-内酰胺类抗生素之前,该产酶菌的耐药性已经存在。日益广泛而大量的β-内酰胺类抗生素的使用,甚至滥用,加速了β-内酰胺酶产酶菌耐药性的形成及复杂化,以致耐药菌株不断增多。其机制除了“持留菌”的因素,还与某些β-内酰胺类抗生素诱导产酶菌基因高突变有关;另外,产β-内酰胺酶敏感菌株通过外源性质粒或(和)转座子的横向传递,接受相关的耐药基因而耐药。β-内酰胺酶耐药基因又可联合其他类抗生素耐药基因编码重组,导致多重耐药菌株产生,并且其耐药水平藉此增强。细菌的耐药水平是多种机制综合作用的结果,这一认识解释了单个抗生素在治疗细菌感染时往往无效甚至容易引起耐药的原因,也是抗生素复合制剂研发的理论依据。细菌的耐药机制从不同的层次和角度看有不同的解释,与“蛋白网络”理论相区别的细菌耐药的经典理论因基于细菌与抗生素相互作用的特异性而被视为“靶标理论”。
“靶标理论”下的细菌耐药机制主要有5种:①生成体外生物膜,阻止抗生素的渗透。②细胞壁外膜孔蛋白减少、缺失或突变,抗生素无法导入菌体。③细胞壁外排系统主动泵出,进入菌体的抗生素达不到杀菌浓度。④青霉素结合蛋白发生改变,以致抗生素不能干扰细胞壁的合成。该机制是革兰阳性菌对β-内酰胺类抗生素耐药的主要原因,提示其与革兰阳性菌的β-内酰胺酶主要是胞外酶有某种相关性。胞外酶是革兰阳性菌分泌至菌体外的β-内酰胺酶,其中也有少量黏附在细胞质膜上。革兰阳性菌中只有葡萄球菌产生β-内酰胺酶。⑤生成灭活或钝化酶类。这其中除了氯霉素乙酰转移酶、氨基苷类抗菌药物钝化酶及大环内酯类抗菌药物钝化酶外,β-内酰胺酶是最具临床意义的,因为临床所见的耐药菌,约80%与β-内酰胺酶的产生有关。产生β-内酰胺酶并通过水解或非水解的方式使β-内酰胺类抗生素灭活则是革兰阴性菌对β-内酰胺酶类抗生素耐药的主要机制,与革兰阴性菌的β-内酰胺酶主要存在于周质间隙成为胞内酶也有某种相关性,其中有少数可通过渗透的方式转移到细胞外:①非水解方式是指抗生素分子到达周质间隙后,β-内酰胺酶与之合成无活性而极其稳定的共价化合物,使之无法作用于青霉素结合蛋白的靶位而失活。相关报道中,该方式较水解方式要少见。②水解方式对于金属β-内酰胺酶而言,其活性位点上的金属离子,通常是Zn2+与相关氨基酸结合导致β-内酰胺环被破坏,进而使抗生素失效。对于丝氨酸β-内酰胺酶而言,这是较多见的方式。其水解过程为:丝氨酸β-内酰胺酶与抗生素结合成非共价键Michaelis复合体,接着该酶活性位点上丝氨酸的自由羟基侵袭β-内酰胺环,生成共价酰酯,酰酯水解使β-内酰胺环开环,经重排后酶失活,同时抗生素自身也被灭活。总之,影响β-内酰胺酶致细菌耐药的因素,就其自身而言,与酶的种类、数量、位置、动力学及生理生化情况等相关;就抗生素而言,与其种类、有效浓度及其与青霉素结合蛋白的亲和力等有关;另外,还与细菌所处的外界环境有关,如温度、酸碱度、诱导剂或培养基等。
主要参考资料
[1] 现代药学名词手册
[2] β-内酰胺酶耐药机制及其中医药相关研究进展
[3] 牛奶中β-内酰胺酶来源及其检测方法研究进展


欢迎您浏览更多关于β-内酰胺酶的相关新闻资讯信息